
- Title: Peptide Probes for Plasmodium falciparum MyoA Tail Interacting Protein (MTIP): Exploring the Druggability of the Malaria Parasite Motor Complex
- Authors: Charlie N. Saunders, Ernesto Cota, Jake Baum, and Edward W. Tate
- Journal: ACS Chemical Biology
- Year: 2020
Malaria is one of the world’s oldest known diseases. In the second century, Romans were advised to wear amulets to protect themselves against the mal aria, or “bad air.” Unsurprisingly, this was ineffective against the surrounding wetlands that were infested with mosquitoes – mosquitoes that we now know carried a Plasmodium parasite. Today, malaria is usually treated with artemisinin or a related molecule along with a second antimalarial drug. This is referred to as artemisinin-based combination therapy. The combination of two different drugs offers the advantage of two different mechanisms of action, which means that the two drugs will have different molecular targets that they interact with to exert their therapeutic effects. Targeting multiple biochemical mechanisms of a disease can help kill the malaria Plasmodium parasite faster, which decreases the risk of the parasite mutating and developing drug resistance.
Even though artemisinin combination therapy is usually effective, resistance to antimalarials is an emerging issue. Researchers are constantly seeking new drug targets, particularly for Plasmodium falciparum, the parasite responsible for most malaria-related deaths. Plasmodium parasites infect red blood cells. The parasite invades cells with the help of a myosin motor, called Myosin A. This motor helps propel the parasite so it can move quickly from cell to cell and gives it the force needed to break through a cell’s membrane. Once in a red blood cell, Plasmodium’s life cycle involves several stages that concludes – and then begins again – with asexual reproduction and bursting of the red blood cell. In malaria and related parasites, this is often referred to as schizogony, because one Plasmodium schizont reproduces to form many discrete merozoites. These merozoites invade other red blood cells with the help of the myosin motor.
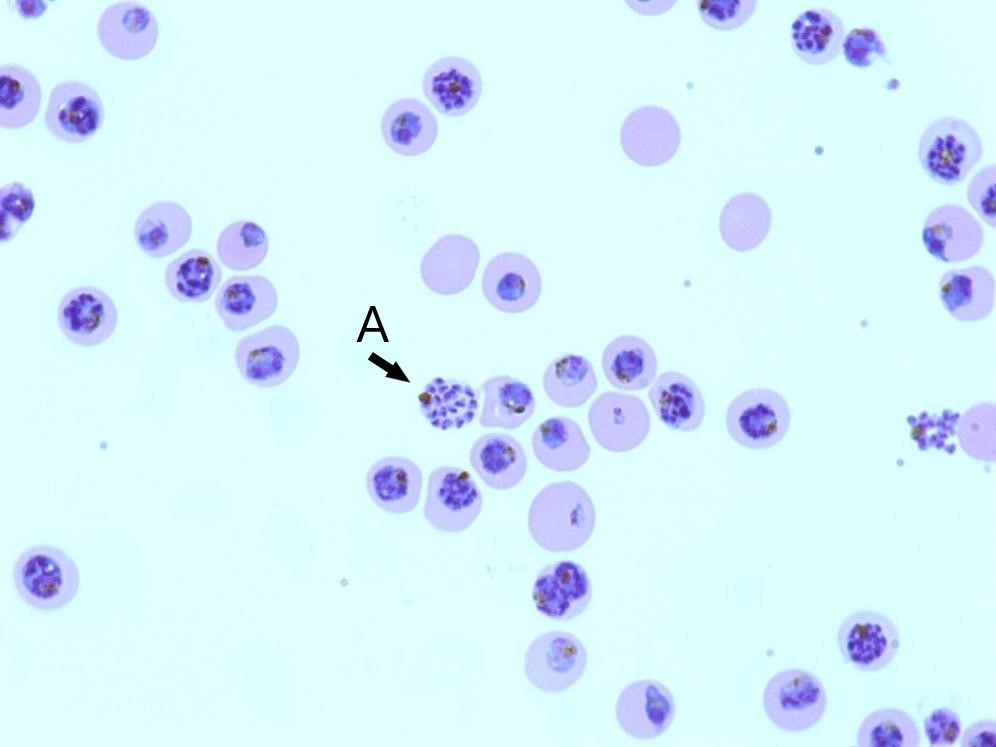
Myosin A has been implicated by researchers as being a potential molecular target for new pharmaceutical agents. If a drug could slow down the parasite by restricting its primary mode of motility, the drug therefore could slow merozoite egress and disease progression. Saunders and coworkers detail their findings in exploring the “druggability” of this target – i.e., exploring the vulnerability of Myosin A activity to therapeutic compounds. One way that researchers may try to “drug” a target is by interrupting a protein-protein interaction (PPI). Myosin A interacts with another protein which helps anchor it. This other protein is called Myosin A tail interacting protein, or MTIP. The PPI between Myosin A and MTIP (written as Myosin A:MTIP) is critical for the motor to function properly. Therefore, this report details attempts to disrupt this important PPI and disrupt motor function.
The researchers noted several key challenges in their attempt to target the Myosin A:MTIP interaction. First, a drug would have to pass through three membranes to reach this target: the red blood cell plasma membrane, the parasitophorous vacuole membrane, and the parasite’s plasma membrane. Second, the aforementioned life cycle of Plasmodium means that the PPI target is only present at certain stages, namely when the parasite is in the schizont or merozoite forms. Therefore, much of the research described in this report details the attempts to make a drug that is amenable to these specific parameters.
The “drug” chosen for optimization, termed My1, was a protein that looks very similar to Myosin A but is shorter and therefore nonfunctional as a motor. This allows for tight binding of the protein to MTIP. The ability of the proteins to interact (also called their binding affinity, or Kd, the equilibrium dissociation constant) was measured via isothermal titration calorimetry. Isothermal titration calorimetry measures the heat absorbed or released when proteins interact. The data is then input into thermodynamic equations (i.e., ΔG = ΔH -TΔS or ΔG = RTlnKd) to determine binding.
After measuring the binding parameters of My1, the researchers began making modifications to make it more suitable for its target. This was done by increasing the positive charge on the protein (i.e., by mutation or addition of amino acids with positively charged side chains) which makes molecules more likely to be taken into cells. The research team also examined several model peptides and found that peptides high in the positively charged amino acid arginine showed promising uptake. Based on these results, My1 was modified to incorporate more arginine. F-ArgMy1, which has five strategically placed arginine residues and a fluorescent molecule (F) attachment, was chosen as the lead candidate because of its increased uptake without too much disruption of the binding affinity. The fluorescent addition offered a way to track and localize the molecule in the cell, since it will emit measurable light under certain conditions. Fluorescent data, along with other experiments, led the team to conclude that F-ArgMy1 does penetrates the red blood cell membrane and may interact with the target protein.
Despite the headway made in these experiments, the modified peptide did not seem to block parasite invasion when tested. Potentially, the therapeutic may not reach exactly where it needs to be within the cell, or perhaps the Myosin A:MTIP interaction is not as vital as was originally thought because of further stabilization by the other proteins that make up the complete parasite motor. This research details the struggles of drugging a specific target and suggests that Myosin A:MTIP is particularly challenging. More work is needed to determine if efforts to drug the myosin motor are indeed fruitless, or if this could be an exciting target for new antimalarials.